va
Results are divided in two main sections:
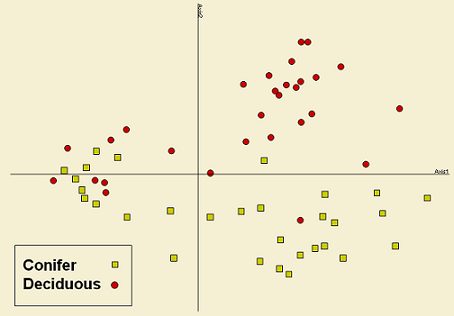
Results from NMS ordination (stress for spiders: 11.27; stress for plants: 18.34) clearly show that forest cover (deciduous vs. coniferous) has a strong influence over both ground-dwelling spiders and vascular plants assemblages (Figure 5). In both graphs almost all sites in conifer forests cluster apart from sites in deciduous forests; thus, Axis2 is describing the variance explained by the type of forest cover in both graphs.
These results are in accordance to those observed for richness and diversity. This is showing that forest cover is an important feature to determine the structure of both communities and that assemblages from both conifer and deciduous stands are different in composition and abundance. Differences in forest structure (e.g. light intensity, temperature, overstory architecture) might be influencing habitat and microhabitat features that are determining the species that are able to survive in these habitats.
In addition, this difference is important at the landscape level. Having a variety of forest cover types and thus different forest successional stages, assures maintaining heterogeneity in the landscape and therefore sustaining biodiversity.
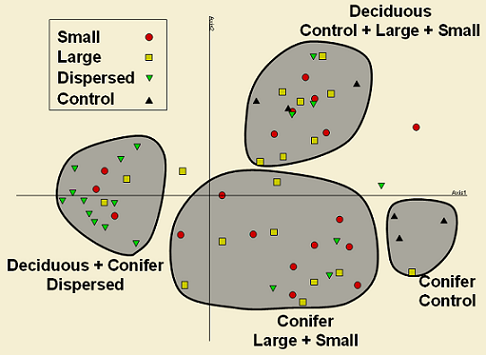
On the other hand, retention class (clumped vs. dispersed) showed a strong effect on the spider assemblage (Figure 6) but not on the plant assemblage (not shown). For clarity in Figure 6, similar sites were surrounded by grey ellipses to show groupings. Thus, dispersed areas of clear-cuts and 10% retention of conifer and deciduous forests cluster distinctively far to the left of the ordination, whereas retained patches were clustered in between dispersed areas (disturbed) and control areas (undisturbed). Although there is no difference between large and small retention patches, it is very interesting that these patches in deciduous forests are very similar in species composition and abundance to controls, whereas in conifer stands, control compartments cluster apart from retention patches.
It has been already shown that some features in the landscape are important to determine and shape ground-dwelling spiders as well as some key environmental factors. The question now is to establish how different assemblages are related to each other and how one is dependent or explained by other. It is widely known that plants are the main drivers for many other species; the forest provides habitats and shapes environmental features that are key for other species.
As a consequence, it is hypothesized that vascular plants in the forest floor are determining habitat and microhabitat features that are shaping and influencing spider assemblages and therefore the observed effects of harvesting on spiders could be explained by the plant assemblage. In other words, that vascular plants in the forest floor could be used to predict spider assemblages associated to the same sites.
This is the purpose of a Co-Correspondence Analysis (CO-CA). As explained in the Data Analysis section, to perform COCA only the most important species in both assemblages were chosen using MRT. Thus, 30 spider species and 48 plant species were chosen running multiple trees removing species from the most rare ones (lower abundance) to the more common (higher abundance) until obtaining a different tree.
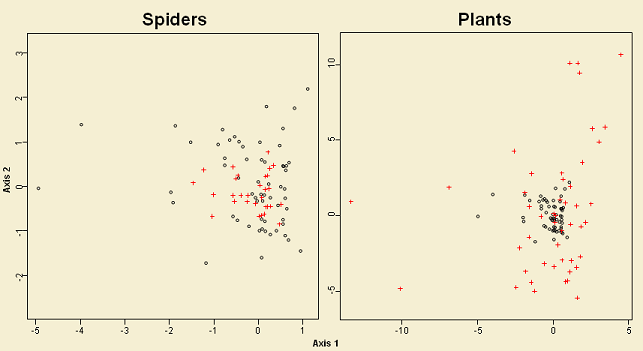
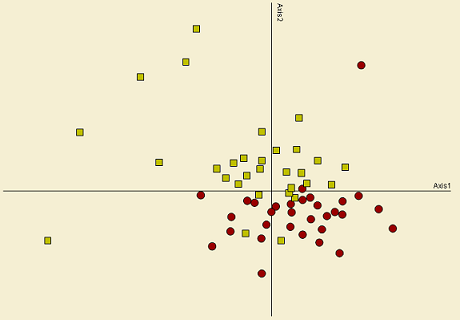
The site pattern (empty circles in Figure 8) in the ordination space for both communities is the same. In addition site distribution due to forest cover, harvest treatment and retention class is similar to the observed on the NMS ordination (e.g. deciduous and conifer sites clustering apart). In contrast, the species pattern (red crosses in Figure 8) for both communities is very different. This is showing that plant assemblages are poor predictors for spider assemblages. About 20% of the variation of spiders is explained by plants, which can be considered very low. As a consequence, the hypothesis outlined above seems to be false; therefore, spider assemblages might be responding to a different group of organisms (e.g. prey items), which in turn might be affected in a similar way by harvesting.
This result suggests that ground-dwelling spiders, and perhaps spiders associated to other forest layers, are responding more to changes in habitat configuration due to disturbance that in turn alters abiotic factors, rather than to species specific assemblages. For example, spiders that are determined by moss cover in conifer forests are being driven not by the species of mosses present in these stands, but by the presence of moss. Those spider species might rely on moisture and ground temperature, for example, rather to the presence of a moss species in the forest floor.
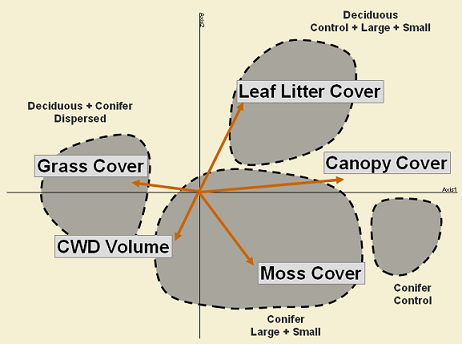
Interestingly, some of these variables are associated to retention patches in the ordination space (Figure 7), suggesting that retained patches are maintaining some features that mimic in some degree undisturbed areas (controls). As a consequence, these patches constitute a very important structural feature in the landscape after human disturbances (harvesting) and are effective for emulating residuals after natural disturbances (fire).
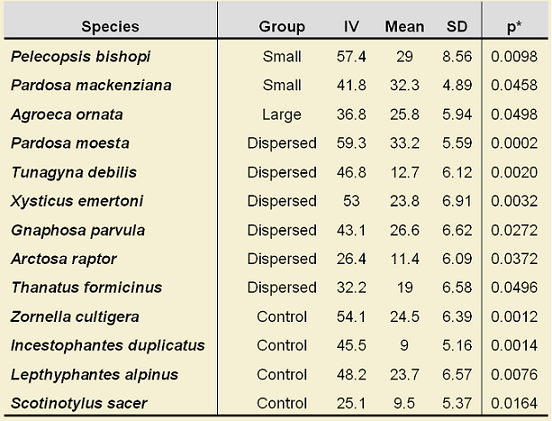
Furthermore, an Indicator Species Analysis (Dufrene & Legendre 1997) was carried out to establish which spider species were more associated to retention classes; thus, 12 species showed a significant (α=0.05) indicator value (IV), although none of these can be considered as strong indicator, since the highest IV was 59% (Table 1). It is interesting to note that all the indicator species for Control areas belong to the family Linyphiidae, while those that are indicator of dispersed retention areas are species commonly found in disturbed areas, such as P. moesta which was not a surprise to be an indicator since, as mentioned before, is probably one of the most abundant and common species after disturbances in the boreal forest. In addition, those species that are indicators for clumped retention (small and large) should be considered as important species; as long as these species are present in retention patches, it might be interpreted that these patches are still functioning as mimics for natural disturbances. Perhaps, increasing the area of these retention patches could improve the number of indicator species and their strength for indication, since larger patches might retain more environmental features that are typical from undisturbed areas.
From the different variables that were measured (click here for more details) some showed to affect species composition and abundance of spiders in both disturbed and undisturbed areas of conifer and deciduous forests. Thus, some environmental variables were strongly associated to clusters in the NMS ordination (Figure 7). As a consequence, leaf litter cover, moss cover, grass cover, canopy cover and CWD volume seem to strongly affect ground-dwelling spider assemblages in the boreal forest.
Although forest cover is one of the most important factors that shapes the ground-dwelling spider community in the boreal forest (Figure 5), environmental variables associated to these habitats are the ones that are driving the community. Thus, deciduous forests are characterized by having a thick litter layer due to the leaf input every year, whereas in the conifer forest floor mosses play an important role. Similarly, undisturbed areas are retaining most of their natural canopy cover that is removed by harvesting, allowing the establishment of open habitat species, such as grasses, which are very common in disturbed areas.
Similarly, NMS showed that there was a minor effect of harvesting treatments on spiders. In the ordination (Figure 5 above), there is a slight differentiation from left to right following a disturbance gradient (not shown for clarity): clear cut - 10% - 75% - Control. In the case of plants, harvesting showed no clustering in the ordination, suggesting that this assemblage is less affected by harvesting. (For a more complete comparison between treatments performing a MRPP, click here).
As noted before, Multivariate Regression Trees (MRT) are a very helpful tool to understand how environmental variables are shaping communities. Thus, it is possible to explore, describe and predict relationships between multiple species and variables (De'ath 2002). After running multiple trees, it became evident that some of the environmental variables that were measured did not help to explain differences in ground-dwelling spider and vascular plant assemblages. However, some of these variables seem to be very important to describe species composition variation across the landscape.
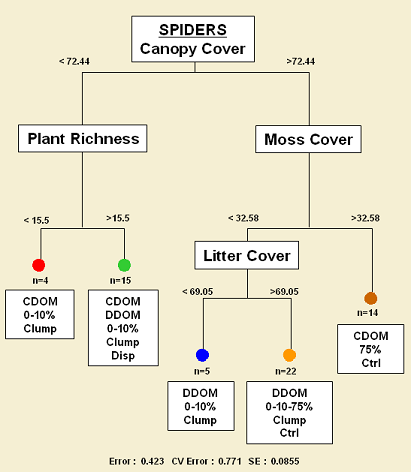
Canopy cover and shrub cover are determining in great extent spider and plant communities (Figure 4). Thus, disturbed areas with canopy cover less than 72% maintain a distinct spider assemblage than less disturbed areas with canopy cover over 72%. For the former sites, plant richness splits the assemblage in two groups whereas for the latter, moss and leaf litter cover determines three groupings. Interestingly, these five branches in the tree are related to the four clusters of sites observed in the NMS ordination (Figures 6 & 7).
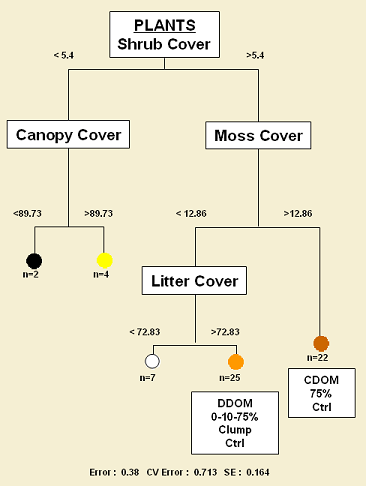
In contrast, for vascular plants in the forest floor, shrub cover might be playing a similar role as canopy cover for spiders; however, most of the sites were grouped (54 of 60) showed values of shrub cover higher than 5%, as a consequence, is difficult to establish if shrub cover actually is an important feature for plants. Nonetheless, as for spiders, moss cover and leaf litter cover are important factors that determine in great extent plant composition in the forest floor of boreal forests.
These areas are dominated by species that are well adapted to disturbances (e.g. wolf spiders), suggesting that retention patches are maintaining some environmental features that prevent in some degree colonization from disturbed areas. In contrast, number of species increase in retention patches, this might be an indication that retention patches act somehow as species refuges and are thus an important feature in the landscape after disturbances, such as harvesting. Perhaps disturbance has a greater effect on spiders in dispersed retention since it might be affecting in a higher degree microhabitat features that are maintained in retention patches; clumped retention, on the other hand, show higher richness values since it is sustaining species from both undisturbed and disturbed conditions.
Both richness and abundance showed different patterns in spiders and plants. For spiders, these parameters were affected by retention class (clumped vs. dispersed) but not by patch size (small vs. large) (Figure 3). A higher species richness was observed in retention patches than in dispersed areas; thus, 40 species were unique in retention patches while 14 were only collected in dispersed areas. It can be observed that patch size affects neither the number of species nor the abundance. Likewise, more individuals were collected in dispersed retention areas in comparison to clumped retained patches. This was somehow expected, given the small difference in area between the small patch (ca 0.15Ha) and the large one (ca 0.4 Ha), allowing operation of similar processes (at least for spiders), such as edge effect.
For plants in contrast, only large retention patches showed higher species richness; in small patches and dispersed areas this parameter was similar and lower (Figure 3). In large patches 11 species were unique, whereas the same number of species were unique in both small patches and dispersed areas.
More highly disturbed areas (clear-cut and 10% retention) account for a greater proportion of overall spider abundance. Specimens from these lower harvest retention treatments comprise 76.44% of the total spiders (clear-cut: 40.67%; 10% retention: 35.77%). In contrast, for plants, harvest treatments seem to have a lower impact on the percentage cover; these highly disturbed areas account for slightly more than half of the total (56.36%; clear-cut: 29.90%, 10% retention: 26.46%), showing a grater proportion alone in the 75% retention treatment (33.38%).
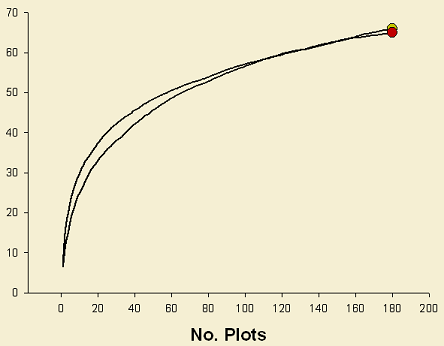
Harvesting treatments seem to have a lower effect on species richness (Figure 2). However, for spiders, under the same number of individuals (ca. 2000 ind.), 75% retention tends to show higher species richness in comparison to 10% retention and clear-cut. Similarly, this same situation is observed for plants. As a consequence these results suggest that there is a relation between richness values and disturbance degree. The higher richness observed in clear-cuts and 10% retention, compared to controls, might be explained by the higher number of open habitat specialists observed in these harvest treatments. As a consequence, 75% retention stands sustain the highest species richness because of the presence of both open and close habitat specialists.
These results show that, on the one hand, conifer forests as late successional stages in the boreal forest are important for maintaining spider biodiversity. Conifer forests sustain higher species richness than deciduous forests, in addition to the high number of species collected on this habitat type. On the other hand, despite the lower richness in deciduous forests, these early successional stages are also important as species refuges after disturbances, maintaining more than half of the species observed in older stands.
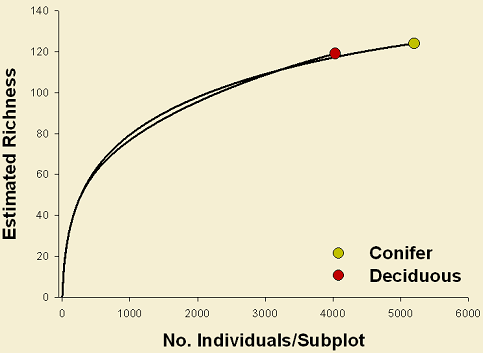
Similarly, there is an effect of cover type on the abundance of spiders and the percentage cover of plants. Higher number of spiders were collected in conifer stands (Figure 1). In contrast, a slight increment in percentage cover was observed in deciduous stands.
Cover type affects in some degree both the ground-dwelling spider and vascular plant assemblages. Although the difference in species richness between conifer and deciduous forests is not evident (Figure 1). For spiders, under 4000 individuals, about 120 species were recorded in both cover types, showing a slight higher richness in deciduous stands. Similarly for plants, about the same number of species were recorded in both cover types. A large proportion of species for both spiders and plants, were shared between conifer and deciduous stands (90 spider species; 53 plant species). However, 63 spider species were unique in either forest type (34 in conifer and 29 in deciduous) and 25 plant species were unique in either forest type (13 in conifer and 12 in deciduous).
Pardosa moesta was the most abundant species (2089 ind.), followed by P. mackenziana (852 ind.), P. uintana (480 ind.), Cybaeopsis eupola (457 ind.), Gnaphosa microps (377 ind.), Alopecosa aculeata (357 ind.) and Diplocentria bidentata (352 ind.). These seven species together account for 53.7% of the total spiders collected. In addition, 91 species were represented by 10 or fewer individuals. Thus, as noted before, the spider assemblage is dominated by wolf spiders, ground spiders and sheet-weavers, 82% of the total number of individuals were represented by these families, specially wolf spiders that account for almost 50%. This huge contribution of this family is due mainly by the high abundance of P. moesta, this species is the most common and abundant in the the boreal forest, specially in disturbed areas(Dondale & Redner 1990, Buddle 2000, Pickavance 2001). In contrast, Linyphiidae (sheet-weavers), alone accounted for more than half of the species recorded.
On the other hand, 78 species of vascular plants were recorded (to go to the plant species list click here). From the total, 12 species were very common and found in more than half of the plots (Cornus canadensis, Rosa acicularis, Epilobium angustifolium, Rubus pubescens, Viburnum edule, Fragaria virginiana, Lathyrus ochroleucus, Petasites palmatus, Linnea borealis, Mertensia paniculata, Mitella nuda and Viola renifolia), whereas 41 species were found in 10 or less plots, accounting for more than 50% of the total. In contrast most of the species that showed the highest average percentage cover were those that were found in few plots (e.g. Equisetum pratense, Salix myrtillifolia).
In total, 180 pitfall traps operated for collecting spiders and 180 plots were displayed for surveying vascular plants. As mentioned in the Data Collection section, these samples were obtained in a combination of 2 forest cover types (conifer/deciduous), three harvesting treatments (clear-cut, 10% & 75% retention) and two retention types (dispersed/clumped retention).
From these samples, 153 species of spiders (15 families and 9234 adult individuals) were recorded (to go to the spider species list click here). As expected, Liniyphiidae, Gnaphosidae and Lycosidae were the most commonly collected spider families in pitfall traps, these families are represented by typical ground-dwelling species such as wolf spiders.
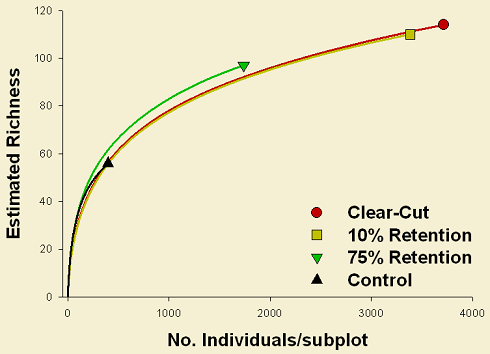
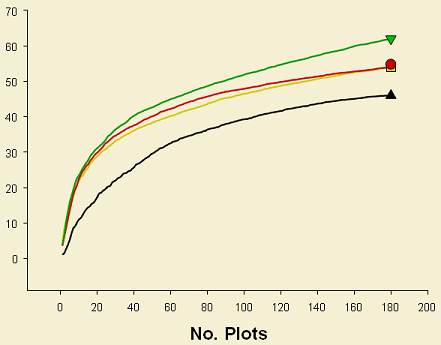
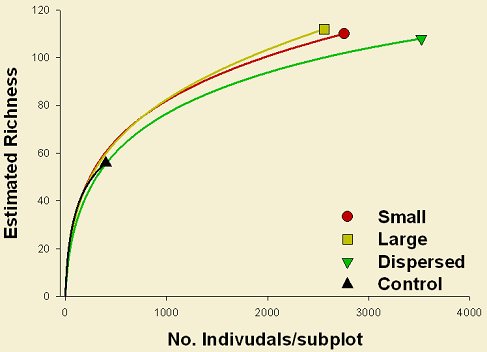
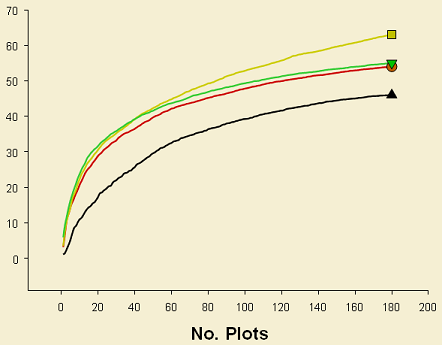
These results are suggesting that retention patches in deciduous forests emulate natural disturbances somehow better than in conifer forests. There is evidence that retention patches are maintaining heterogeneity for ground-dwelling spiders, which is lost in dispersed retention areas (all samples cluster tightly) and that these patches are actually functioning as species refuges after disturbances, being more similar to control areas.