J Pharm Pharmaceut Sci (www.ualberta.ca/~csps) 8(1):1-9, 2005
Evaluation of the first-pass glucuronidation of selected flavones in gut by Caco-2 monolayer model.
Siu Pong Ng, Ka Yin Wong, Li Zhang, Zhong Zuo1
School of Pharmacy, Faculty of Medicine, The Chinese University of Hong Kong, Shatin, New Territories, Hong KongGe Lin
Department of Pharmacology, Faculty of Medicine, The Chinese University of Hong Kong, Shatin, New Territories, Hong KongReceived 27 September 2004, Revised 20 November 2004, Accepted 22 November 2004, Published 20 December 2004
PDF Version
Abstract
PURPOSE. Four flavones, namely Apigenin, Baicalein, Chrysin and Luteolin, were selected for study and comparison of their absorption and metabolism in gut using the in vitro Caco-2 monolayer model. METHODS. Transport of the four flavones in the Caco-2 monolayer model was studied in both Apical-to-Basolateral and Basolateral-to-Apical directions. RESULTS. All of the selected flavones were able to pass through the Caco-2 cell monolayer with no significant efflux. The permeability coefficients of the four compounds were all greater than 10-6 cm/sec and those of Apigenin and Baicalein were even greater than 10-5 cm/sec. Glucuronides of the tested flavones were all formed in the Caco-2 cell monolayer model and a structure-activity relationship has been proposed for this glucuronidation. In addition, Apical-to-Basolateral transport studies were performed in Caco-2 models pre-treated with Chrysin, an UGT inducer. Quantities of the corresponding glucuronides formed were all significantly higher in Chrysin-treated groups than the controls. CONCLUSIONS. It demonstrated that all selected flavones were substrates of the UGT isoforms that are inducible by Chrysin.
Introduction
Flavonoids, classified mainly into four subgroups: flavone, flavonol, flavanone, isoflavone (1), are polyphenolic compounds that usually exist in plants as secondary metabolites. They possess strong antioxidative activity (2) as well as other potential beneficial effects including anti-atherosclerotic (3), anti-inflammatory (4), anticancer (5), anti-thrombogenic (6), antiviral (7) and anti-osteoporotic effects (8).
In general, flavonoids occur in plants either in the form of aglycones or glycosides. The aglycones are capable of being absorbed freely from gut by passive diffusion, while the flavonoid glycosides are usually hydrolyzed to the corresponding aglycone molecules prior to its gastrointestinal absorption (9). Nevertheless, both as aglycone or glycosides, the flavonoid bioavailabilities have not been found as high as expected from their favorable lipophilicity. Such discrepancy is believed to be mainly due to their extensive first-pass metabolic effect in small intestine (10).
Phase II metabolisms such as glucuronidation or sulfation seem to be the major metabolic pathway for flavonoids (11). The importance of the first-pass effect exerted by the intestine is generally regarded to be less than the liver due to the lower protein level and catalytic activity of the drug metabolizing enzymes in gut (12). However, Chrysin, a member of the flavone subgroup, had been demonstrated to have very low oral bioavailability in the healthy volunteers due to its extensive first-pass metabolism, mainly glucuronidation in gut (13). Moreover, substrates prone to glucuronides may be pumped out of the enterocytes back to the lumen by the organic anion efflux transporter, for example, the multidrug resistance associated protein 2 (MRP2) (14). Such a mechanism is proposed to be the main reason for the poor bioavailability of Chrysin in humans, based on the findings using the in vitro Caco-2 cell monolayer model (13, 15).
Among all the available in vitro systems for the study of mucosal drug absorption and metabolism, the Caco-2 cell model is one of the most popular models utilized to rapidly assess the cellular permeability of potential drug candidates (16). Typical small-intestinal microvillus hydrolases and nutrient transporters can be found in the Caco-2 cell monolayer model. In addition, this model could also allow the study of presystemic drug metabolism in gut since differentiated Caco-2 cells also express various cytochrome P450 isoforms and Phase II enzymes such as UDP-glucuronosyltransferases, sulfotransferases and glutathione-S-transferases (17).
Therefore, four flavone aglycones namely Chrysin (C); Apigenin (A), Baicalein (B) and Luteolin (L) (Figure 1), respectively, were selected for the current study to 1) investigate their transport and metabolism mechanism, namely the Phase II glucuronidation, across the intestinal mucosa; 2) evaluate the possible UGT isoforms responsible for the glucuronidation of the selected flavones in the in vitro Caco-2 cell monolayer model; and 3) establish a possible structure-activity relationship for the first-pass metabolism of the selected flavonoids in small intestine.
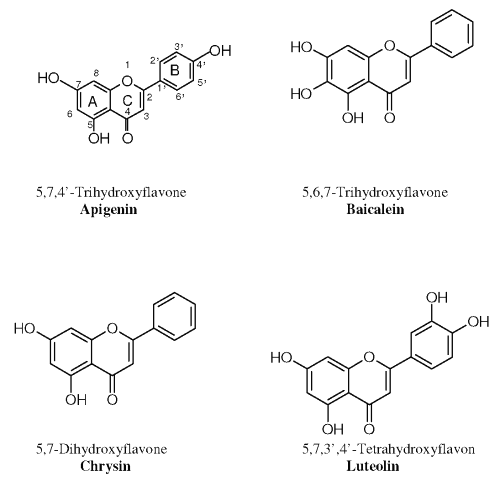
Figure 1: Structures of the four selected flavones.
Materials and Methods
Materials
Apigenin, b-glucuronidase and phosphate buffered saline tablets were purchased from Sigma Chem. Co. Baicalein was obtained from Aldrich Chem. Co. Chrysin and trifluoroacetic acids (TFA) were purchased from Sigma-Aldrich. Luteolin was purchased form Lancaster Synthesis. HPLC grade methanol was from Fisher Chemicals. Analytical grade acetone and dimethyl sulfoxide (DMSO) were from Lab-Scan Analytical Sciences. Sodium dihydrogenphosphate monohydrate, disodium hydrogen orthophosphate (AnalaR® grade) and acetic acid were from BDH Laboratory Supplies.
The Caco-2 cell line (passage 18) was obtained from American Type Culture Collection (ATCC). Dulbecco's modified Eagle's medium, fetal bovine serum, nonessential amino acid, L-glutamine, sodium pyruvate and penicillin/streptomycin used in cell culture were all supplied by GibcoBRL®. The Transwell® inserts (sterile, Polycarbonate filters) were purchased from Corning Costar Corporation.
A Reichert Scientific-Temperature Controlled Water Shaker was used in solubility tests, stability test and sample hydrolysis treatment. A Waters 2695 Separations Module equipped with a Waters 996 Photodiode Array Detector was employed in the HPLC analysis. Labconco Centrivap Concentrator and Eppendorf Centrifuges 5415 D & 5810R, were also used.
Methods
Preparation of standard and buffer solutions
Stock solutions of selected flavone. Stock solutions (4 mg/mL) of each selected flavones were prepared by dissolving accurately weighted appropriate amount of the chemicals in DMSO. Stock solutions of each selected flavone (40 mM) for preparation of HPLC calibration curves was then prepared by further dilution of their 4mg/mL stock solutions with methanol. All solutions were vortexed, sonicated and then stored at -20°C for further uses.
Phosphate buffered saline plus (PBS+). The phosphate buffered saline plus (PBS+) solution was used as transport buffer in the Caco-2 monolayer model. It was prepared by dissolving a phosphate buffered saline tablet in 200 mL of deionized water, followed by the addition of 90 ml of 2 M calcium chloride and 80 ml of 1 M magnesium chloride.
HPLC analysis
The HPLC analysis were performed with a reversed-phase column (100 x 4.6mm 5 m Hypersil® ODS C18, Jones Chromatography) equipped with a guard column (7.5 x 4.6mm 5 m Spherisorb® ODS-2 C18, Waters), running with an isocratic mobile phase of 55 % methanol in 0.3 % TFA at a flow rate of 0.9 mL/min (18).
Preparation of calibration curves
Solutions for calibration curves were prepared by mixing the appropriate amount of standard stock solutions of each selected flavones with pH 2.5 phosphate buffer to reach the final concentrations of 0.1, 0.2, 0.5, 1, 1.5, and 2 μM for each compound, respectively. A 100 ìL aliquot of the prepared standard solution was mixed with 100mL of internal standard in methanol:0.3% TFA (50:50,v/v). The internal standard (I.S.) used for Apigenin, Baicalein and Chrysin was 0.5 mM of Luteolin and that used for Luteolin was 1 mM of Apigenin. The mixture (100 mL) was then injected into HPLC system for analysis.
Solubility test of the selected flavones in PBS+ at 37°C
Excess amount of the flavones was added to 10 mL of PBS+ and the mixture was then vortexed and sonicated. Three sets of mixture were put into a water bath at 37°C for 24 h, and subsequently, the mixtures were taken out and centrifuged. The supernatant was then passed through a 0.45 mm cellulose acetate filter. The last mL of filtrate collected was diluted appropriately by phosphate buffer pH 2.5 and a 100 mL aliquot was mixed with 100 mL of the corresponding I.S. for HPLC analysis. The average concentration calculated is regarded as the solubility at 37°C.
Stability test of the selected flavones in PBS+ of pH 7.4 at 37°C
Each flavone (40 mM) was prepared with PBS+ at pH 7.4 and kept at 37ºC water bath. Samples were taken at pre-determined time points (0, 30, 60, 90, 120, 240 & 360 min) followed by proper dilution with phosphate buffer of pH 2.5. All the tests were done in triplicate. The amount of each compound remained in PBS+ was determined by HPLC and the residue amount of each compound was plotted against time to obtain their stability profiles.
Caco-2 Cell Culture
Caco-2 cells were cultured in Dulbecco's modified Eagle's medium supplemented with 10% fetal bovine serum, 1% nonessential amino acid, 1% L-glutamine, sodium pyruvate and penicillin/streptomycin. Cells were first grown in humidified atmosphere (90% relative humidity) with 5% CO2 at 37ºC in 75 cm2 culture flask, and then were sub-cultured after 4 days using 0.05% trypsin-0.53mM EDTA.
Caco-2 cells were seeded at a density of 3x105 cells/well in Transwell® inserts (24 mm, 0.4 mm pore size, 4.71 cm2, Polycarbonate filter) previously coated with a thin collagen layer. The medium was changed every other day for a total of 21 days. The integrity of the Caco-2 monolayer was monitored by the transepithelial electrical resistance (TEER) for the whole period. The transport experiments were carried out at 20-23 day after seeding. The cells from passage 39 to passage 43 were used in the current study.
Transport and metabolism studies of the selected flavones in Caco-2 monolayer model
Verification of the concentrations for the loading solutions. A loading solution (40 mM) of each flavone was prepared by adding appropriate volume of their 4mg/mL stock solutions in PBS+ of pH 7.4. The prepared solutions were then passed through a 0.45 m m cellulose acetate filter and diluted with phosphate buffer pH 2.5. Three sets of tests were conducted for each flavone. The concentration of the filtrates was verified by HPLC analysis.
Bi-directional transport of the selected flavones. Following the removal of culture medium and pre-equilibration with PBS+ at 37°C for 15 min, 1.5/2.6 mL of the 40 mM loading solution for each selected flavone was added to the donor side (Apical/Basolateral of the Transwell® insert). The sample (0.5 mL) was then taken from the receptor side (Basolateral or Apical) at different time intervals of 15, 30, 45, 60, 90 and 120 min. An equal volume of blank PBS+ was replaced to the receptor side immediately after each sampling. Sample was also withdrawn from the donor side at 120 min. The percentage of each compound recovered in forms of both parent drug and metabolites from both apical and basalateral sides was calculated as Recovery%.
Calculation of apparent permeability coefficient (P app ) for transport studies. P app , expressed in cm/sec, was calculated by the previously described equation (19).
P app = (dc/dt×V)/(A×C o ) where

Data points used in the calculation of P app should be under sink condition (e.g., the receptor side contains not more than 10 % of initial loading dose of drug in donor side).
Transport and metabolism of selected flavones in UGT inducer-treated Caco-2 monolayer model
Pretreatment of UGT inducer (Chrysin) on the Caco-2 monolyer model. Pretreatment of the Caco-2 cells seeded in the Transwell® with 50 mM of Chrysin was conducted 48 h prior to the transport experiment (20). Chrysin was dissolved in ethanol:DMSO (80:20, v/v) at a final concentration of DMSO not exceeding 0.5 %. The culture medium was changed every 24 h.
Effect of the pretreatment of Chrysin on cell integrity. Tests were performed to assess whether the 48 h Chrysin treatment would alter the integrity of the monolayer or not. Transwells® seeded with Caco-2 cells would undergo the following treatments: 1) treated with Chrysin as described in 2.2.8.1; 2) treated with the same volume of blank solvent used in treatment 1) (ethanol:DMSO, 80:20, v/v) for the same period of time; 3) untreated, to serve as a negative control.
A 40 mg/mL aliquot of propranolol was loaded to the apical side of the Transwell®. Samples were taken at various time intervals of 15, 30, 60, 90, 120 min for HPLC analysis by UV at 291 nm. Papp values of propranolol obtained from different treatments were then calculated and compared.
Apical-to-Basolateral transport studies in Chrysin-treated Caco-2 monolayer models
A 40 mM aliquot of the loading solution of each selected flavone was added to the apical side of the Transwell® with and without Chrysin treatment. Samples were taken from the basolateral side at 30, 60 and 120 min, and from the apical side at 120 min.
Sample analysis
Samples from the above transport studies were tested for the concentration of both the parent compound and its glucuronides. The amount of glucuronides in the samples was determined by hydrolysis treatment with b-glucuronidases. The difference in amount of flavone aglycone before and after the enzymatic hydrolysis treatment was regarded as the amount of flavonoid glucuronides present in the samples.
Sample hydrolysis and extraction. A 100 mL aliquot of sample was added with 10 mL of 2 M sodium acetate buffer (pH 5.0) and 25 mL of b -glucuronidase solution (12500 m/mL) freshly prepared with 0.2 M sodium acetate buffer (pH 5.0) (21). After incubation at 37°C for 45 min, 50 mL of internal standard in methanol was added into the above mixture. For Apigenin, Baicalein and Chrysin, 1 mM of Luteolin was used as internal standard, while 2 mM of Apigenin was used as internal standard for Luteolin.
The mixture was then extracted once with 1 mL of acetone by vortex mixing and then centrifuged at 4,000 rpm for 5 min. The supernatant was taken and evaporated to dryness by the centrifugal concentrator at room temperature. The residue was reconstituted with 200 mL of mobile phase. After centrifugation at 13,200 rpm for 10 min, 100 mL of the supernatant was injected into HPLC for analysis.
Preparation of calibration curves for samples with extraction treatment. Apigenin, Baicalein, Chrysin and Luteolin standard solutions in PBS+ were prepared with final concentrations of 0.2, 0.5, 1, 1.5, 2 and 4 mM. These solutions were treated following the hydrolysis and extraction procedure mentioned above. Triplicates were conducted for each concentration. The mean peak area ratio of the studied flavone versus internal standard was plotted against concentration of the flavone to obtain the standard curves.
Percentage of recovery from sample hydrolysis and extraction procedure. The absolute extraction recovery for hydrolysis and extraction procedure was determined by comparing the peak area of known concentration of the flavones (0.2 & 4 mM) before and after the process.
Statistical analysis
Student's unpaired t-test was used to evaluate the statistical differences in concentrations of the flavones or their glucuronides between the different treatment groups. Differences were considered significant when the P-value calculated was less than 5 %. Values are expressed as the mean ± standard error of mean (SEM).
Results
Sample analysis of the selected flavones
All calibration curves of flavones with authentic compounds were found to be linear with correlation coefficients greater than 0.999. At the lowest concentration of 0.1 mM, all selected flavones produced peaks at least 3 times greater than that of the background.
All the calibration curves for Apigenin, Baicalein, Chrysin and Luteolin after the hydrolysis and extraction treatment were also linear with correlation coefficients greater than 0.995. The absolute extraction recoveries for the four flavones were within the range of 73 to 88 %.
Solubility and stability test
Among the four selected flavones, Apigenin is the least soluble with a solubility of 3.27 ± 0.06 mM at 37°C in PBS+, while Luteoline the most soluble (59.95 ± 1.89 mM). Solubilities of Chrysin and Baicalein were 4.34 ± 0.12 mM and 27.42 ± 0.21 mM, respectively. Stability profiles of the four compounds at pH 7.4 37°C are shown in Figure 2.
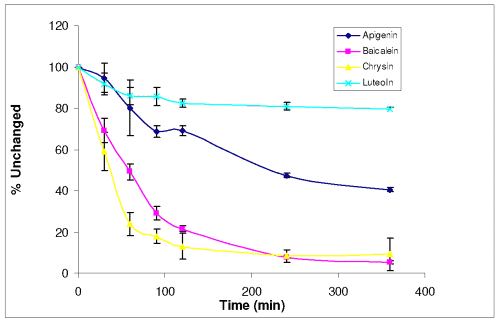
Figure 2: Stability profiles of Apigenin, Baicalein, Chrysin and Luteolin (40 mM) (n=3).
There were 40.46 %, 5.24 %, 9.14 % and 79.35 % of Apigenin, Baicalein, Chrysin and Luteolin remained in PBS of pH 7.4 after 6 hours, respectively.
Transport and metabolic studies of the selected flavones in Caco-2 monolayer model
Verification of the loading solution concentrations. The actual concentration of the 40 m M loading solutions for each flavone were found to be: 32.33 ± 0.78 mM for Apigenin, 37.77 ± 0.96 mM for Baicalein, 35.05 ± 1.03 mM for Chrysin and 36.52 ± 0.85 mM for Luteolin. Therefore, all flavones dissolved well in PBS+ at 37°C.
Papp values from the bi-directional transport of the selected flavones. The Papp values of all the selected flavones obtained from both Apical to Basalateral and Basalateral to Apical directions are shown in Table 1.
Table 1: Data on bi-directional transport study of the selected flavones (n=3).
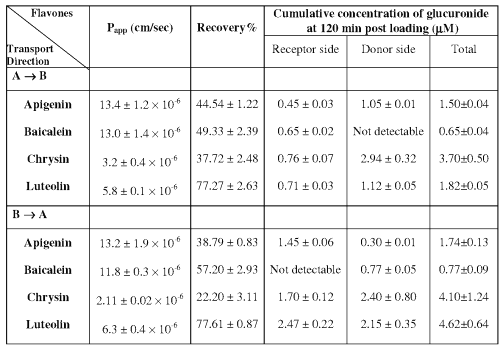
The P app values of Apigenin and Baicalein from both directions are about 5- and 3- fold higher than that of Chrysin and Luteolin. In addition, there are no directional transport differences for all the studied flavones, indicating that efflux may not exist during the transport of these compounds. The percentages of the flavones recovered from both the donor and receiver side of the Caco-2 monolayer model at the end of the 120 min incubation period are also presented in Table 1. It was observed that the percentage recovery of each flavone was much greater than what have been shown from the stability study.
Formation of glucuronide during the transport of the selected flavones. All of the selected flavones underwent glucuronidation during their bi-directional transport. The cumulative concentrations of glucuronide at both donor and receptor side at 120 min post loading are also shown in Table 1. Regardless of the side of the loading, Baicalein glucuronide was barely detectable at the end of the transport study in the apical side. In general, the amount of glucuronide formed from the Basalateral to Apical direction is similar to that from the Apical to Basalateral direction. Glucuronides formed in both donor and receptor sides were more pronounced in Chrysin and Luteolin, whereas the lowest glucuronide level was generated in Baicalein.
Transport and metabolism of selected flavones in the UGT inducer-treated Caco-2 monolayer model
Effect of pretreatment of Chrysin on cell integrity. The Papp values of propranolol calculated for the first 30 min in Chrysin-treated, solvent-treated and untreated Caco-2 were 15.21 ± 3.68 × 10-6 , 18.72 ± 2.48 × 10-6 and 23.28 ± 1.91 × 10-6 cm/sec respectively, with no significant difference from each other (P > 0.05). This suggests that the permeability of the cell monolayer is not likely altered by Chrysin or organic solvent treatment as applied in our study.
Effect of Chrysin on the glucuronide formation of the selected flavones. As shown in Figure 3, the cumulative glucuronide concentrations of the selected flavones found on the receptor side of the Chrysin-treated models were all significantly higher than those from the control, through the whole incubation period.
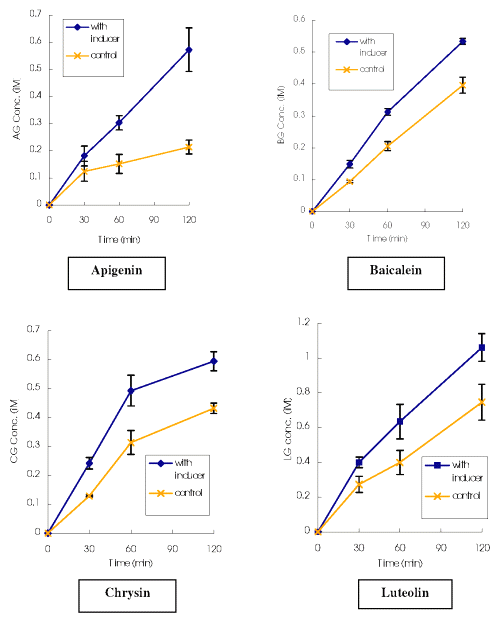
Figure 3: Cumulative glucuronide concentrations of the selected flavones (followed Apical to Basalateral transport in Caco-2 model) versus time in Transwells® with and without UGT inducer treatment. AG: Apigenin; BG: Baicalein; CG: Chrysin; LG: Luteolin; * Significantly greater than control (p<0.05). ** Significantly greater than control (p<0.01). *** Significantly greater than control (p<0.001).
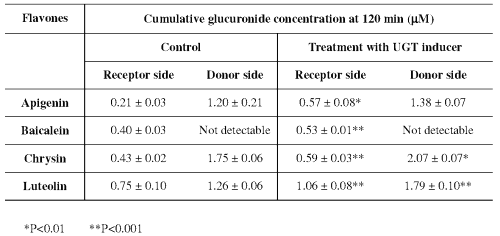
It was also observed (Table 2) that the amounts of glucuronide generated on the donor side of Chrysin and Luteolin are also significantly higher in the UGT inducer-treated model than the control at 120 min, whereas this trend towards increase was not obvious for Apigenin and Baicalein.
Table 2: Effect of a UGT inducer on the formation of glucuronide of flavones in the Caco-2 cell (n=3).
Discussion
The purpose of this study was to gather information on transport and metabolism of the selected flavonoids from the same subgroup of flavone. Significant amounts of glucuronides of all tested flavonoids were found in the Caco-2 cell monolayer model. On the basis of these observations in the Caco-2 model, we postulate that glucuronidation could be a major metabolic pathway for flavonoids in the gut wall. Physiochemical tests of solubility and stability performed in the current study not only support these data for flavonoids in the in-vitro model, but also provide information on flavonoid properties that may affect their absorption.
Solubility and stability of selected flavonoids. The four selected flavonoid aglycones were all poorly soluble in aqueous buffer solution with solubilities below 20 mg/mL at 37°C. Thus, one of the primary causes for the poor bioavailability of flavones, such as Chrysin, may be the poor solubility.
Therefore, a co-solvent system was employed for the loading solution for the Caco-2 model, as aqueous buffer by itself was not capable of producing a loading concentration that was high enough to give samples detectable by HPLC. Thus, less than 0.3% DMSO was added to the loading solution in our study in order to improve the solubility as well as to avoid the damage to the cell monolayer (22).
Although the poor stability of Chrysin and Baicalein in pH 7.4 PBS + was found from the in vitro incubation tests with around 7% of Chrysin and 10% of Baicalein left after 2 hour, the percentage of Baicalein and Chrysin that remained in the Caco-2 model (with over 50% for Baicalein and 35% for Chrysin) was much higher than that observed in the in vitro stability test. The presence of Caco-2 cells may thus improve the stability of the flavonoids, which is consistent with the observations from Crespy et al (23). The overall absorption of the chemically unstable compounds, such as Baicalein and Chrysin, in Caco-2 monolayer model could be a competition among several processes including chemical degradation of the compounds, permeation of the compounds across the cell monolayer, uptake of the compounds into the Caco-2 cells, metabolism of the compounds inside the Caco-2 cells, excretion of the metabolites that formed within the Caco-2 cells. Compared with Apigenin and Luteolin, the still relatively low recoveries of Baicalein and Chrysin from Caco-2 cell model could also potentially due to their relative high uptake by the Caco-2 cells, different metabolic rate and excretion rate of their formed metabolites, which may need further proof.
A possible structure-stability relationship has been identified for the studied compounds in PBS+ of pH 7.4. As shown in Figure 1, Apigenin and Luteolin, possessing at least one hydroxyl group in the B ring, are much more stable than Baicalein and Chrysin, which do not have any substituents on the B ring. Opening of the C-ring through oxidation is usually a major degradation pathway of 4-oxo flavonoids such as quercetin, one of the most well studied flavonoids (24). It was proposed that flavonoids with substituents that could protect the C-ring from opening would be much more stable in aqueous solution. Thus, the hydroxyl groups on the B ring of Apigenin and Luteolin are considered to be able to protect the C-ring from opening through the formation of resonance with that on the C-ring.
Transport and glucuronidation of selected flavones in Caco-2 monolayer models. The Papp values of Apigenin and Baicalein fell into the range of 10-5 cm/sec, which implies that they would be well absorbed in gut due to their high lipophilicities (25). The Papp values of the 4 flavones obtained from both directional transport studies were similar. Our calculated Papp from apical to basalateral for Chrysin differs a bit from the observation by Walle et al (15), which may result from the differences in Caco-2 cell passage number, cell culture condition, transport medium and the use of co-solvent in the loading solution (22). However, our observation for Luteolin differs from that obtained by Tammela (26) et al, in which Luteolin showed no apical to basalateral transport. This could be due to the different detection capability of Luteolin from transport study using different loading concentrations, different size of the transwell insert.
Previous studies from both Apigenin and Chrysin have shown that they could be extensively metabolized in Caco-2 cell model and their hydrophilic phase II conjugates including glucuronide and sulfate could be excreted to apical side by MRP and OATs (15, 27). The selective localization of Baicalein glucuronides from our study suggests that there must be active transporter involved also. In general, MRP2 and MRP3, found in the apical and basolateral sides of the intestine epithelium (28), are responsible for the excretion of conjugated metabolites to the lumen and blood supplies, respectively (14). Similar to the gut, Caco-2 cells also express MRP2 and MRP3 (28). Although involvement of other transporters may also occur, organic anions including conjugative metabolites are generally considered be poor substrates for P-glycoprotein (14). The absence of Baicalein glucuronide on the apical side may imply that it is a much stronger substrate of transporters located at the basolateral side, such as MRP3, rather than the substrate of the transporters on the apical side, i.e. the MRP2.
The total amount of flavonoid glucuronides found on both sides of the Cao-2 cell model, in descending order, was Chrysin>Luteolin>>Apigenin>>Baicalein. It was noticed that conjugation would most likely happen at the 7-OH position of the selected flavones due to the intramolecular hydrogen binding on 5-OH position. Therefore, the higher amount of glucuronide of Chrysin and Luteolin could possibly due to their favored nucleophilicity at the 7-OH position.
Transport and metabolism of flavonoids in the UGT inducer-treated Caco-2 monolayer model. The amount of glucuronides secreted into the basolateral side was significantly higher in the Chrysin-treated models than that in the control for all the selected flavones. It is therefore possible that all of the 4 selected flavone can be metabolized by the UGT isoforms inducible by Chrysin, most likely the UGT1A1, not UGT1A6, 1A9 or 2B7 isoforms (20). However, the UGT isoforms UGT1A8 and 1A10, which are also expressed in the intestine (13) and capable of catalyzing the glucuronidation of flavonoids (30), have not shown to be inducible by Chrysin. Thus, further study is needed to verify the specific isoforms involved in the glucuronidation of the selected compounds.
Our study not only providing more information on the mechanisms of flavonoid absorption but also addresses the potential oral absorption problem for herbal products. Although various in vitro studies have demonstrated potential beneficial effects for all sorts of flavonoids, flavonoids may not be effective in vivo due to either their low solubility or the high degree of first-pass metabolism. The above results emphasized the important role of gut wall first-pass metabolism of flavonoid-containing herbal products after oral intake. In addition, the data generated from flavonoids with similar structures would provide information on the structure-bioavailability relationships for those with poor bioavailability.
Acknowledgement
CUHK Direct Grants (CUHK 2040830, CUHK 2041012).
References
Phenols and phenolic glycosides. In: Evans WC. Trease & Evans’ Pharmacognosy. 14th ed. WB Saunders London, pp 250-251, 1996.
Rice-Evans, CA. Miller, NJ. and Paganga, G., Structure-antioxidant activity relationships of flavonoids and phenolic acids. Free Radic Biol & Med, 20:933-956, 1996.
Arai, Y., Watanabe, S., Kimira, M., Shimoi, K., Mochizuki, R. and Kinae, N., Dietary intakes of flavonols, flavones and isoflavones by Japanese women and the inverse correlation between quercetin intake and plasma LDL cholesterol concentration. J Nutr, 130:2243-2250, 2000.
Lin, JK., Tsai, SH. and Lin, SY., Anti-inflammatory and anti-tumor effects of flavonoids and flavanoids. Drugs of the Future, 26:145-152, 2001.
Birt, DF., Hendrich, S. and Wang, W., Dietary agents in cancer prevention: flavonoids and isoflavonoids. Pharmacol Ther, 90:157-177, 2001.
Tzeng, SH., Ko, WC., Ko, FN. and Teng, CM., Inhibition of platelet aggregation by some flavonoids. Thromb Res, 64:91-100, 1991.
Bae, EA., Han, MJ., Lee, M., Kim, DH., In vitro inhibitory effect of some flavonoids on rotavirus infectivity. Biol Pharm Bull, 12:1122-1124, 2000.
Hegarty, VM., May, HM., Khaw, KT., Tea drinking and bone mineral density in older women. Am J Clin Nutr, 71:1003-1007, 2000.
Manach, C., Regerat, F., Texier, O., Agullo, G., Demigne, C. and Remesy, C., Bioavailability, metabolism and physiological impact of 4-oxo-flavonoids. Nutr Res, 16:517-544, 1996.
Pietta P, Gardana C, Mauri PL., Identification of Gingko biloba flavonol metabolites after oral administration to humans. J. Chromatogr. A, 693:249-255, 1997.
Kuhnau, J., The flavonoids, a class of semi-essential food components: their role in human nutrition. World Rev Nutr Diet, 24:117-191, 1976.
Lin, JH., Chiba, M. and Baillie, TA., Is the role of the small intestine in the first-pass metabolism overemphasized? Pharmacol Rev, 51:135-157, 1999.
Tukey, RH. and Strassburg, CP., Human UDP-glucuronosyltransferases: Metabolism, expression, and disease. Annu Rev Pharmacol Toxicol, 40:581-616, 2000.
Suzuki, H. and Sugiyama, Y., Role of metabolic enzymes and efflux transporters in the absorption of drugs from the small intestine. Eur J Pharm Sci, 12:3-12, 2000.
Walle, UK., Galijatovic, A., Walle, T., Transport of flavonoid Chrysin and its conjugated metabolites by human intestinal cell line Caco-2. Biochem Pharmacol, 58:431-438, 1999.
Hidalgo, IJ., Cultured Intestinal Epithelial Cell. In: Borchardt RT, Smith PL, Wilson G. Models for Assessing Drug Absorption & Metabolism. Plenum Press, New York: pp 35-50, 1996.
Meunier, V., Bourrie, M., Berger, Y. and Fabre, G., The human intestinal epithelial cell line Caco-2; pharmacological and pharmacokinetic applications. Cell Biol. Toxicol, 11:187-194, 1995.
Galijatovic, A., Walle, UK. and Walle, T., Induction of UDP-Glucuronosyl-Transferase by flavonoids Chrysin and quercetin in Caco-2 cells. Pharm Res, 17:21-26, 2000.
Artursson, P., Epithelial transport of drugs in cell culture. I: A model for studying the passive diffusion of drugs over intestinal absorptive (Caco-2) cells. J Pharm Sci, 79:476-482, 1990.
Galijatovic, A., Otake, Y., Walle, UK. and Walle, T., Induction of UDP-glucuronosyltransferase UGT1A1 by flavonoid Chrysin in Caso-2 cells-potential role in carcinogen bioinactivation. Pharm Res, 18:374-379, 2001.
Erlund, I., Kosonen, T., Alfthan, G., Maenpaa, J., Perttunen, K., Kenraali, J., Parantainen, J. and Aro, A., Pharmacokinetics of quercetin from quercetin aglycone and rutin in healthy volunteers. European Journal of Clinical Pharmacolog, 56:545-53, 2000.
Yamashita, S., Furubayashi, T., Kataoka, M., Sakane, T., Sezaki, H. and Tokuda, H., Optimized conditions for prediction of intestinal drug permeability using Caco-2 cells. Eur J Pharm Sci, 10:195-204, 2000.
Crespy, V., Morand, C., Besson, C., Manach, C., Demigne, C. and Remesy, C., Comparison of the intestinal absorption of quercetin, phloretin and their glucosides in rats. J Nutri, 131: 2109-2114, 2001.
Boulton, DW., Walle, UK. and Walle, T., Fate of the flavonoid quercetin in human cell lines: Chemical instability and metabolism. J Pharm Pharmacol, 51:353-359, 1999.
Artursson, P. and Karlsson, J., Correlation between oral drug absorption in humans and apparent drug permeability coefficients in human intestinal epithelial (Caco-2) cells. Biochem. Biophys. Res Commun, 175:880-885, 1991
Tammela, P., Laitinen, L., Galkin, A., Wennberg, T., Heczko, R., Vuorela, H., Slotte, JP. and Vuorela, P., Permeability characteristics and membrane affinity of flavonoids and alkyl gallates in Caco-2 cells and in phospholipid vesicles. Arch Biochem Biophys, 425:193-9, 2004.
Hu, M., Chen, J. and Lin, H., Metabolism of flavonoids via enteric recycling: mechanistic studies of disposition of apigenin in the Caco-2 cell culture model. J Pharmacol Exp Ther 307:314-321, 2003.
Borst, P., Evers, R., Kool, M. and Wijnholds, J., The multidrug resistance protein family. Biochim. Biophys Acta, 146:347-357, 1999.
Hirohashi, T., Suzuki, H., Chu, XY., Tamai, I., Tsuji, A. and Sugiyama, Y., Function and expression of multidrug resistance-associated protein family in human colon adrenocarcinoma cells (Caco-2). J Pharmacol Exp Ther, 292:265-270, 2000.
Cheng, Z., Radominska-Pandya, A. and Tephly, TR., Studies on the substrate specificity of human intestinal UDP-glucuronosyltransferases 1A8 and 1A10. Drug Metab Dispos, 27:1165-1170, 1999.
Corresponding Authors: Zhong Zuo, Room 610, Basic Medical Science Building, School of Pharmacy, The Chinese University of Hong Kong, Shatin, N.T. Hong Kong. joanzuo@cuhk.edu.hk
Published by the Canadian Society for Pharmaceutical Sciences.
Copyright © 1998 by the Canadian Society for Pharmaceutical Sciences.
http://www.cspscanada.org